
Intensywny trening aktywuje syntezę białek, ale tylko wtedy, gdy istnieje odpowiednie odżywianie, które ją wspiera. Jeśli czytasz artykuł, który napisaliśmy z Johnem Meadowsem, zatytułowany Maksymalizacja syntezy białek, lub jeśli czytałeś cokolwiek, co T Nation mówi na ten temat od lat, prawdopodobnie znasz pojęcie „okna anabolicznego” i znaczenie odżywiania okołotreningowego.
To, co dzieje się na poziomie komórkowym w ciągu kilku godzin po treningu, zapowiada długoterminowe zyski. Skorzystaj z tego „okna anabolicznego”, a będziesz rosnąć jak nigdy dotąd. Konsekwentnie za tym tęsknisz i, cóż, powodzenia.
Kluczem jest uzyskanie właściwych makroskładników we właściwym czasie, ale makra to tylko część całościowego obrazu. Ważnym, ale często pomijanym aspektem syntezy białek mięśniowych jest objętość komórek. Objętość komórek to nie tylko kosmetyka; jest głównym motorem transportu aminokwasów, działając również za kulisami, aby włączyć syntezę białek i powstrzymać ich degradację.
Mięsień pełny / o zwiększonej objętości to mięsień anaboliczny. Chociaż od ponad 20 lat wiemy, że obrzęk komórek hamuje rozpad białek i stymuluje syntezę białek w niektórych komórkach (1-3), do niedawna mechanizm łączący objętość komórki z syntezą białek był tajemnicą.
Teraz wiemy, że synteza białek jest kontrolowana przez enzym mTOR, który jest aktywowany przez stres mechaniczny, czynniki wzrostu i leucynę.
Chociaż wszystkie trzy z nich są ważne dla bodźca treningowego, sygnalizacja mTOR zależy również od objętości komórek.(4) Jest to szczególnie ważne w mięśniach szkieletowych, gdzie wolumizacja komórek aktywuje syntezę glikogenu i białek oraz hamuje ich rozpad.(5, 6)
Przełom naukowy prowadzący do powiązania objętości komórek z syntezą białek nastąpił w 2005 roku, kiedy to grupa naukowców odkryła, że do aktywacji mTOR potrzeba czegoś więcej niż tylko leucyny - potrzebna jest również glutamina.(7)
To była niespodzianka. Chociaż glutamina jest uważana za „warunkowo niezbędny” aminokwas, który ogranicza rozpad białek podczas poważnych urazów lub stresu, nigdy nie była powiązana z aktywacją mTOR.
Glutamina była niezbędna do wchłaniania leucyny i zwiększania objętości komórek, z których oba są potrzebne do włączenia syntezy białek. Autorzy następnie pokazali tę komórkową glutaminę wyczerpanie nie tylko powoduje zmniejszenie objętości komórek, ale także zmniejsza zdolność leucyny do aktywacji syntezy białek.(7)
To odkrycie było ogromne, ponieważ zapewniło bezpośredni związek między glutaminą, wolumizacją komórek i syntezą białek. Po raz pierwszy wykazano, że glutamina jest niezbędna do aktywacji syntezy białek przez leucynę.
Chociaż to badanie sugeruje, że glutamina jest bardzo ważnym elementem układanki łączącej objętość komórek i syntezę białek, dokładny mechanizm został opracowany dopiero w 2009 roku, kiedy Nicklin i wsp. odkryli, że eksport glutaminy jest powiązany z importem leucyny i aktywacją mTOR.(8)
Aby leucyna dostała się do komórki, następuje początkowy okres „ładowania glutaminy”.„To również wciąga wodę, zwiększając objętość komórek. Po fazie „ładowania glutaminy” glutamina jest eksportowana z komórki w zamian za import leucyny.
Nicklin i in. odkryli również, że komórkowe poziomy glutaminy ograniczają szybkość aktywacji syntezy białek przez leucynę. Gdy komórki były jednocześnie traktowane glutaminą i mieszaniną EAA zawierającą leucynę, aktywacja syntezy białek była opóźniona o 60 minut. Kiedy te same komórki były „wstępnie załadowane” glutaminą, synteza białek została włączona w ciągu 1-2 minut po podaniu leucyny do komórek.
Wynik ten był ważny, ponieważ wyjaśnił opóźnienie aktywacji syntezy białek przez leucynę w tym modelu eksperymentalnym.
Wyniki te w końcu rzuciły trochę światła na maszynerię komórkową, która reguluje transport aminokwasów i jak jest ona połączona z kontrolą syntezy białek.
Jednak tę pracę należy interpretować z pewną ostrożnością. Ważnym zastrzeżeniem do tych badań było to, że przeprowadzono je in vitro (tj.mi. w kulturach komórkowych), gdzie regulacja lub synteza białek jest znacznie prostsza. Komórki mięśniowe są zdolne do wytwarzania glutaminy w razie potrzeby z innych aminokwasów, a „niedobór glutaminy” w tym modelu hodowli komórkowej nie jest reprezentatywny dla bardziej fizjologicznych sytuacji in vivo.
Wysokie tempo syntezy białek nie może być utrzymywane w nieskończoność w tkance mięśniowej z suplementacją glutaminy lub bez niej. Glutamina może być jednak stosowana do strategicznego wspomagania syntezy białek poprzez optymalizację objętości komórek w okresie potreningowym.

Komórki są bardzo zajęte i istnieje wiele związanych z błoną kanałów jonowych i białek transportowych, które regulują ruch do iz komórki. Szczególnie ważne są tu dwie klasy transporterów aminokwasów: transportery aminokwasów „System L” i „System A” są najściślej powiązane z sygnalizacją mTOR i syntezą białek.(8–10)
Aktywność transporterów Systemu A i Systemu L jest sprzężona, co umożliwia wchłanianie leucyny i innych BCAA do komórki.(11) Transportery Systemu L są odpowiedzialne za napływ leucyny i innych BCAA w zamian za wypływ glutaminy.
Transportery systemu A działają jednak według innego mechanizmu, w którym glutamina jest sprzężona z poborem sodu.(12, 13) Sprzężenie między poborem sodu a transporterami aminokwasów Układu L / Układu A nazywa się Tertiary Active Transport (w skrócie TAT). To TAT ostatecznie kieruje leucynę do wnętrza komórki, prowadząc do aktywacji mTOR i syntezy białek.(11)
Jak działa TAT, możesz zobaczyć na poniższym rysunku:
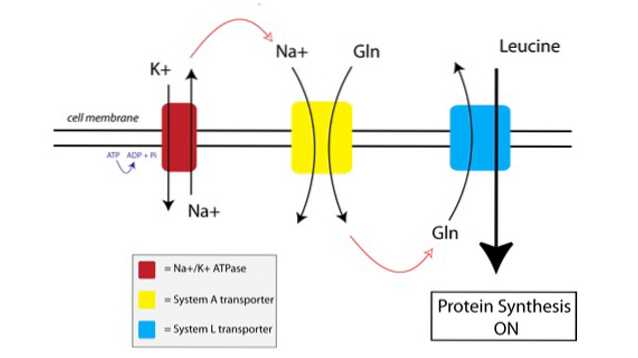
Po pierwsze, pompa membranowa zwana pompą sodowo-potasową ATPazy (Na + / K + ATPaza, czerwona na powyższym rysunku) wykorzystuje energię ATP do przemieszczania sodu poza komórkę, wbrew gradientowi stężenia.
Zwiększone stężenie sodu na zewnątrz komórki jest sprzężone z importem glutaminy przez transporter Systemu A (kolor żółty na rysunku). Napływ glutaminy i sodu do komórki również przyciąga dodatkową wodę, powodując pęcznienie komórki. To wprowadza komórkę w stan anaboliczny, przygotowując mechanizm syntezy białek do aktywacji.
Kiedy glutamina gromadzi się w komórce do dostatecznie wysokiego poziomu, aktywowane są transportery Systemu L (niebieskie na rysunku), które przemieszczają glutaminę poza komórkę w zamian za pobór leucyny. Wniknięcie leucyny do komórki jest wyzwalaczem syntezy białek.
Chociaż jak dotąd była to świetna lekcja biochemii, odkrycie TAT jest ważne nie tylko dla biologów komórkowych. Teraz, gdy wiemy, jak objętość komórek jest sprzężona z transportem aminokwasów i syntezą białek, możemy zaprojektować kilka strategii żywieniowych, aby zmaksymalizować proces, gdy ma on znaczenie - w krytycznym okresie po treningu.
Synteza białek jest całkowicie zależna od nawodnienia komórek - jeśli jesteś nawet trochę odwodniony, zdolność regeneracji po intensywnym treningu jest całkowicie ograniczona. Zdobycie dużej ilości wody jest tutaj oczywiste, ale sama woda nie wystarczy.
Elektrolity, takie jak sód, potas, chlorek i fosforan, również pełnią funkcję „osmolitów”, ponieważ wciągają wodę do komórki. Po intensywnym treningu potrzebujemy wody, aminokwasów i elektrolitów, aby zmaksymalizować proces wolumizacji komórek, który napędza syntezę białek.
Sód, magnez, wapń, potas, fosforan i chlorek (żeby wymienić tylko kilka) są tutaj ważne. Na minimalnym poziomie (chyba że robisz to zgodnie z zaleceniami lekarza), nie unikaj sodu przed lub po treningu. Jeśli masz niedobór sodu, Twoja pompa podczas treningu prawie nie będzie istnieć, a sód jest niezbędny do wchłaniania glutaminy.
Aby wyeliminować zgadywanie, Surge® Workout Fuel i Plazma ™ zostały zaprojektowane z idealnymi proporcjami elektrolitów, aby wspierać objętość komórek i syntezę białek.
Wychwyt glutaminy do komórki powoduje wolumizację komórek, przygotowując komórki mięśniowe do syntezy białek. Jak wspomniano, mięsień o pełnej objętości / objętości to mięsień anaboliczny. Wraz z napędzaniem transportu aminokwasów, wolumizacja komórek zwiększa również syntezę glikogenu i hamuje rozpad białek.(4-6)
Synteza białek jest hamowana przez niedobór glutaminy, co ma ogromne konsekwencje dla ciężko trenujących sportowców. Po intensywnym treningu dochodzi do reakcji zapalnej, która pozwala komórkom odpornościowym przejść do pobitej tkanki mięśniowej, aby rozpocząć proces naprawy / odbudowy.(14)
Glutamina jest tak szybko wchłaniana przez komórki odpornościowe, że jest uważana za „paliwo układu odpornościowego.”(15) Nic dziwnego, że wykazano, że intensywny trening powoduje wyczerpanie glutaminy w osoczu.(16–18)
Z tego powodu zapotrzebowanie na glutaminę wzrasta w okresie potreningowym, gdzie lokalna odpowiedź immunologiczna może konkurować o dostępność glutaminy do pobudzenia komórek mięśniowych do wychwytu aminokwasów i syntezy białek.
Ładowanie wstępne komórki z glutaminą mogą również potencjalnie skrócić „czas opóźnienia” związany z aktywacją leucyny w syntezie białek. Jeśli jeszcze tego nie robisz, weź 10-15g glutaminy lub peptydów glutaminy natychmiast po treningu. Ponieważ BCAA to kolejny ulubiony substrat do syntezy glutaminy w mięśniach, a także wykazano, że zwiększają produkcję glutaminy w mięśniach (19-21), BCAA i leucyna są również przydatne w okresie przedtreningowym, aby pomóc zmaksymalizować produkcję endogennej glutaminy.
Od redakcji: Cały protokół Plamza / MAG-10, wraz z odżywianiem przedtreningowym, treningowym i potreningowym, spełnia zalecenia autora dotyczące ładowania mięśni BCAA i leucyną.
Niedawno odkryto, że spożywanie EAA zwiększa ekspresję transporterów aminokwasów zarówno z Systemu A, jak i Systemu L.(9) Co ważne, dzieje się to na „poziomie potranskrypcyjnym”, czyli na poziomie syntezy białek, gdzie istniejące mRNA ulegają translacji na białka.
Porównaj to z „denovo”Ekspresja białek - gdzie synteza, przetwarzanie i transport nowych mRNA może zająć 16 lub więcej godzin - potranskrypcyjna aktywacja syntezy białek może nastąpić w ciągu minut do godzin, umożliwiając komórkom szybkie zwiększenie poziomu poszczególnych białek w razie potrzeby.
Nagle mamy większą motywację do posiadania solidnego planu żywieniowego przedtreningowego - spożycie EAA w okresach przed i okołotreningowych opłaca się dużym potreningiem, zwiększając ekspresję transporterów aminokwasów, przygotowując komórkę do maksymalnego wchłaniania aminokwasów i aktywacja syntezy białek.
Idealne są tutaj szybko wchłaniające się izolaty lub hydrolizaty białek, takie jak Mag-10® Protein Pulsing Protocol ™ lub Plazma ™ w okresie przedtreningowym i okołotreningowym.
Insulina to najbardziej anaboliczny hormon w organizmie. Wraz z bezpośrednią aktywacją syntezy białek insulina zwiększa również translokację transporterów aminokwasów z Układu A do błony komórkowej.(22)
Oznacza to, że insulina powoduje wyświetlenie większej liczby transporterów Systemu A na błonie komórkowej, gotowych do wprowadzenia większej ilości glutaminy do komórki. Więcej glutaminy prowadzi do większej objętości komórek, co prowadzi do większej ilości leucyny do komórki, ostatecznie prowadząc do większej syntezy białek.
Podczas gdy EAA zwiększają wyrażenie transporterów AA to sygnał insuliny, który pozwala im wyświetlać się na powierzchni komórki, gotowe do transportu nowych aminokwasów do komórki.
To kolejny powód, dla którego węglowodany przed i po treningu są dobrym pomysłem, chyba że jesteś w trybie ekstremalnej utraty tłuszczu: insulina zwiększa zdolność transportu aminokwasów komórkowych.
Węglowodany zwiększają poziom insuliny, ale niektóre aminokwasy mogą być również używane do zwiększania wydzielania insuliny. Glutamina jest silnym aktywatorem hormonów „inkretyn”, które powodują, że komórki trzustki produkujące insulinę są bardziej wrażliwe na glukozę.(23) Glicyna nasila również uwalnianie insuliny poprzez inny mechanizm.
Chociaż same węglowodany potreningowe zwiększą poziom insuliny, połączenie tych wzmacniających insulinę aminokwasów z węglowodanami doładuje trzustkę, aby uwolnić jeszcze więcej insuliny. Chociaż dobrze jest przez większość czasu utrzymywać poziom insuliny na niższym poziomie, zwiększony poziom insuliny w okresie okołotreningowym maksymalizuje transport aminokwasów, objętość komórek i syntezę białek, jednocześnie hamując rozpad białek.
Rodzaj intensywnego treningu beztlenowego potrzebny do zbudowania wysokiej jakości mięśni prowadzi do znacznej produkcji mleczanu, obniżając pH mięśni. Prowadzi to do wczesnego zmęczenia i osłabienia mięśni, ale również niektóre transportery aminokwasów, w tym System A, są hamowane przez niskie pH.(13)
Kiedy pH mięśni jest niskie, wychwyt aminokwasów jest zmniejszony, co hamuje aktywację syntezy białek przez mTOR.(24) Wykazano również, że niskie pH hamowanie transporterów aminokwasów z Układu A zwiększa rozpad białek.(25)
Tutaj pojawia się beta-alanina. Podwyższony poziom karnozyny w mięśniach działa jak naturalny bufor kwasowy, wydłużając próg beztlenowy poprzez ograniczenie spadku pH mięśni podczas treningu.
Beta-alanina ma jeszcze jedną ważną funkcję: pomaga utrzymać syntezę białek i przyspieszyć jej działanie online po intensywnym treningu, zapobiegając osłabieniu transportu aminokwasów.
Aby zwiększyć poziom karnozyny w mięśniach, przyjmuj dwie tabletki Beta-7 ™ trzy razy dziennie.
Podczas intensywnych sesji treningowych synteza białek jest zmniejszona, a degradacja białek jest aktywowana. Jest to nieuniknione dla każdego ciężko trenującego podnośnika. Jednak stopień, w jakim możemy zminimalizować kataboliczne skutki treningu i szybciej możemy powrócić do „trybu anabolicznego” w okresie po treningu ostatecznie decyduje o tym, jak skutecznie odzyskamy siły - i będziemy rosnąć.
Czas na spożycie makroskładników jest ważny, ale jest środkiem do osiągnięcia celu. Objętość komórek jest głównym motorem transportu aminokwasów i syntezy białek. Dzięki zrozumieniu, w jaki sposób zachodzi transport aminokwasów i jak jest regulowany przez objętość komórki, możemy szybciej dostarczyć więcej leucyny do zniszczonych komórek mięśniowych, podsycając w ten sposób anaboliczny ogień i ostatecznie prowadząc do lepszych przyrostów.
Powyższe strategie są skuteczne, praktyczne i oparte na najnowszych badaniach naukowych. Użyj ich jako szablonu, aby przenieść swoje postępy w treningu na wyższy poziom.
Czekam na Twoje pytania na LiveSpill!



Jeszcze bez komentarzy